3.2. Sistemas aeróbicos de producción energética
Este proceso proporciona de forma continua átomos de hidrógeno procedentes del catabolismo de los hidratos de carbono, grasas y proteínas almacenados. La mitocondria dispone de mecanismos capaces de extraer electrones a estos hidrógenos ( Oxidación) y transferir finalmente el oxígeno que hemos obtenido del aire atmosférico ( Reducción).
El metabolismo muscular puede ser aerobico y anaerobico. El musculo decide que tipo de sistema utilizar dependiendo de los diversos factores, entre los que sobresale la intensidad del ejercicio ( la reposición de ATP).

La sistematización de los sistemas energéticos en función del tipo de sustrato utilizado, podemos determinar las siguientes clasificaciones:
- Metabolismo de Fosfágenos
- Metabolismo de los Hidrato de Carbono
- Metabolismo de las Grasas
- Metabolismo de las Proteínas
Metabolismo de los Fosfágenos
El metabolismo de los fosfágenos o los fosfatos de alta energía proporciona la energía necesaria para la contracción muscular al inicio de la actividad y durante el ejercicios explosivos, muy cortos y de elevada intensidad. Por ejemplo, en las contracciones isométricas del cuádriceps al 50-75% de la fuerza máxima voluntaria, solo hay ATP suficiente para mantenerla durante cinco segundos, y el Pcr proporciona otros 14 segundos de contracción. La glucólisis comienza a proveer energía desde el quinto segundo.
Metabolismo de los Hidratos de Carbono
Los hidratos de carbono son necesarios en nuestro organismo . La Glucosa nos permite el acceso de obtener energía tanto en condiciones aeróbicas como en condiciones anaeróbicas, Es el único sustrato que la célula utiliza para obtener energía con o sin la presencia de oxígeno.
El proceso por el cual las células obtiene energía de la glucosa en condiciones anaeróbicas se le llama glucolisis. El producto final de este proceso dentro de la célula es la producción de ácido láctico ( o de lactato, ya que realmente la constante de disociación del ácido láctico {pk} este hace que se disocie inmediatamente en el medio interno en un ion lactato y un hidrogenion).
La obtención de energía a partir de la glucosa en condiciones aeróbicas engloba diversos procesos metabólicos celulares, que implican a la actividad mitocondrial. Inicialmente la glucosa experimenta la reacciones iniciales de la glucólisis anaeróbica, si bien el paso final en el que el piruvato se transforma en lactato no tiene lugar . En su lugar, el piruvato se introduce en la mitocondria y tras sufrir una transformación se incorpora a el ciclo de Krebs.

El proceso por el cual las células obtiene energía de la glucosa en condiciones anaeróbicas se le llama glucolisis. El producto final de este proceso dentro de la célula es la producción de ácido láctico ( o de lactato, ya que realmente la constante de disociación del ácido láctico {pk} este hace que se disocie inmediatamente en el medio interno en un ion lactato y un hidrogenion).
La obtención de energía a partir de la glucosa en condiciones aeróbicas engloba diversos procesos metabólicos celulares, que implican a la actividad mitocondrial. Inicialmente la glucosa experimenta la reacciones iniciales de la glucólisis anaeróbica, si bien el paso final en el que el piruvato se transforma en lactato no tiene lugar . En su lugar, el piruvato se introduce en la mitocondria y tras sufrir una transformación se incorpora a el ciclo de Krebs.

Metabolismo de las Grasas
Las grasas se encuentran almacenados en :

3.2.3. Gluconeogénesis y oxidación del lactato durante el ejercicio

Gluconeogénesis:

Las grasas se encuentran almacenados en :
- Musculo
- tejido subcutáneo en forma de gotas de triglicéridos
Representan una reserva casi inacabable de energía, algunas células como las nerviosas no pueden utilizarlas como fuentes energética y dependen exclusivamente de la glucosa. Las grasas producen de un 40 a un 45% de las calorías totales. El empleo de las grasas para la obtención de la energía es tan importante como el de Hc.
Gran parte de los Hc ingeridos se transforman en triglicéridos, se almacenan y se emplean posteriormente para obtener energía. El proceso de utilización de los lípidos es lento.
Está integrado por una serie de pasos :
- Movilización: Ruptura de los triglicéridos almacenados en el tejido adiposo.
- Circulación: Transporte de los ácidos grasos al musculo conjuntamente con la albumina.
- Captación: Entrada de los ácidos grasos libres al musculo.
- Activación: Aumento del grado energético de los ácidos grasos antes de catabolizarlos
- Traslación: Entrada de los ácidos grasos a las mitocondrias
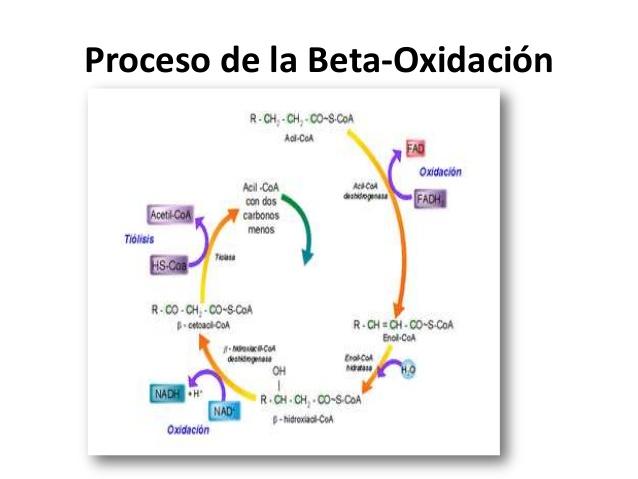
- Beta-Oxidación: Catabolismo de los ácidos grasos a acetil-CoA y producción de hidrogeniones unidos al NAD +(nicotidamina) y FAD ( dinucleótido de adenina flavina )
- Oxidación Mitocondrial: Ciclo de Krebs y cadenas respiratorias.
Metabolismo de las Proteínas
La función de las proteínas es estructural, enzimática o contractil pero en ciertas ocasiones también puede proporcionar energía. Podrían cubrir del 5-10% del gasto energético total de las actividades de larga duración. Para proporcionar energía el aminoácido debe primero convertirse en una forma que pueda acceder fácilmente a las vías de liberación energética.
3.2.1. Oxidación de hidratos de carbono durante el ejercicio
La función de las proteínas es estructural, enzimática o contractil pero en ciertas ocasiones también puede proporcionar energía. Podrían cubrir del 5-10% del gasto energético total de las actividades de larga duración. Para proporcionar energía el aminoácido debe primero convertirse en una forma que pueda acceder fácilmente a las vías de liberación energética.
3.2.1. Oxidación de hidratos de carbono durante el ejercicio
Oxidación de Carbohidratos:
El NADH formado en el citoplasma por efecto del proceso glicolítico poder ser reoxidado en la membrana mitocondrial, el piruvato abandona el citosol y entra en la mitocondria para completar su metabolismo hasta transformarse en H2O y CO2 con la presencia de oxígeno, conservando también el redox citosólico y por vía aeróbica la obtención de energía a partir de la glucosa. A lo anterior se le denomina Glucólisis aeróbica, donde se transforma de glucosa a piruvato o glucólisis, la transformación del piruvato en acetil-CoA, la entrada de este en el Ciclo de Krebs para su degradación y la fosforilación oxidativa.
Las fases de Ciclo de Krebs y fosforilación oxidativa son comunes para el metabolismo energético de lípidos y de proteínas, ya que el acetil-CoA y los elementos intermedios del ciclo de Krebs proceden de otras fuentes energéticas además del piruvato. El piruvato se convierte en acetil-CoA atravesando la membrana mitocondrial por el complejo enzimático piruvato deshidrogenasa (PDH) en la membrana interna mitocondrial. Durante el ejercicio, la elevación de los niveles de adrenalina plasmática provoca la activación de PDH, dando una mayor utilización del metabolismo de los carbohidratos.
En el Ciclo de Krebs, una vez obtenido el acetil-CoA la porción acetil se incorpora al oxalacetato, pasando por acciones enzimáticas que lo volverán a convertir en oxalacetato. En este proceso cíclico se produce la degradación del acetil y la extracción de los H+ (reduciendo NAD y FAD) y los átomos de carbono en forma de CO2, además se produce una transferencia de energía directa donde se fosforila un GDP para transformarse en GTP (guanosín trifosfato).
La fosforilación oxidativa se refiere a los procesos que se acoplan al interior de la mitocondria, por el cual se incorpora un grupo fosfato al ADP para resintetizar el ATP, siendo un proceso endergónico que utiliza la energía liberada en la oxidación. El transporte de electrones se realiza en la membrana interna mitocondrial y debido a ella se lleva a cabo el proceso de la oxidación, donde los H+ liberados en la glucólisis, la transformación de piruvato en acetil-CoA y el Ciclo de Krebs, así como otro proceso como la B-oxidación en grasas, son transportados por coenzimas celulares como el NAD y el FAD hasta el interior de la matriz mitocondrial; allí los H+ se liberan descomponiéndose a su vez en H+ y un e-. El último compuesto de la cadena respiratoria es el citocromo A3, el cual transfiere los electrones al oxígeno molecular obtenido del aire atmosférico, transformándose en oxígeno iónico, el cual se une al H+ para formar H2O y toda la energía liberada en este proceso es utilizada para refosforilar el ADP.
Oxidación de los lípidos:
Sin toma en cuenta de donde vienen los ácidos grasos, antes de ser oxidados y dentro del interior del miocito experimentan un proceso de activación para elevar su nivel energético para ceder energía. Tiene lugar en el sarcoplasma, la unión de una coenzima A (CoA) al ácido graso para dar acil-CoA, este según la necesidad celular puede esterificarse y almacenarse en forma de triglicéridos en la célula muscular u oxidarse. El acil-CoA para oxidarse debe entra a la mitocondria y los ácidos grasos en las fibras tipo I u oxidativas. La carnitina procede de dos fuentes: de la síntesis endógena por el hígado (75%) y la dieta, actuando en la membrana mitocondrial transportando grupos acilo de cadena corta, media y larga a través de ella, encontrada en la membrana interna y externa de esta; las acil-carnitin transferasas y la carnitina permiten el paso exclusivo del radical acil sin que pierda su nivel energético, al atravesar la membrana interna mitocondrial se unen de nuevo a un CoA, dando en conclusión que los ácidos grasos son preferiblemente oxidados en la fibras tipo I. En el interior de la mitocondria los acil-CoA se someten a la B-oxidación, el cual es un proceso metabólico que oxida el radical acilo en el carbono B, dando lugar de nuevo a un acil-CoA con 2 átomos de carbono más un acetil-CoA. Durante este proceso se produce la extracción de átomos de hidrógeno unidos a sus electrones, y en cada B-oxidación se reduce un NAD y un FAD (dando NADH y FADI), si por ejemplo 18 átomos de carbono en su radical acilo experimenta 8 veces la B-oxidación, reduciendo 8 NAD y FAD y dando lugar a 9 moléculas de acetil-CoA. El acetil-CoA entra en el Ciclo de Krebs al igual que procedente del piruvato, la oxidación de los carbohidratos, la fosforilación oxidativa donde irán los hidrógenos del Ciclo de Krebs más los transportados por el NADH y FADH reducido en la B-oxidación.
3.2.3. Gluconeogénesis y oxidación del lactato durante el ejercicio
Oxidación
del lactato:
Esta la opción de
reutilizar el lactato como combustible en otras células musculares diferentes
que lo han producido, en este caso las fibras glagolíticas tipo II. Las fibras
tipo I y los miocitos cardíacos son captadores de lactato sanguíneo que va a
ser utilizado como combustible transformado en ácido pirúvico e introduciéndolo
en la mitocondria para su posterior oxidación. En reposo el lactato en sangre
es de un 50% mientras que en el ejercicio este aumenta hasta un 75-80% (Brooks,
2002), siendo este el principal destino del lactato en la recuperación. Otro
agente importante es la adrenalina, en la contracción muscular por estimulación
de receptores beta adrenérgicos y el consumo neto de lactato de este mismo
(Hamann, 2001). En otras células musculares donde se realiza la recuperación
activa (ejercicio de intensidad moderada o baja, en lugar del reposo) es donde
el mecanismo de oxidación pude ser más acentuado, redistribuyendo los
carbohidratos del músculo.

Es la síntesis de
glucosa nueva a partir de otros metabolitos y es necesaria como fuente de
energía si en caso fuese necesaria al momento en que la glucosa disminuye por
falta de alimento o cuando este es utilizado en el ejercicio de alta
intensidad. Órganos como el cerebro que dependen de glucosa como combustible
primario o los Eritrocitos que la utilizan como único combustible. Los
principales sustratos de la gluconeogénesis son el glicerol, el lactato, la
alanina y el piruvato, pero en el caso respectivo del ejercicio es principalmente
el lactato al ser una fuente predominante de átomo de carbono y que además este
se lleva a cabo en el músculo, específicamente en el periodo de recuperación
después del ejercicio. La conversión de lactato en glucógeno muscular es más
importante en las fibras glicolíticas (de tipo II) que en las oxidativas de
contracción lenta (de tipo I), la cuales oxidan lactato. Los estudios de
Bangsbo y cols. Indican que en la primera hora de recuperación la reconversión
de lactato a glucógeno constituye entre un 13 y un 16% de la resíntesis total
de glucógeno después de ejercicios breves de elevada intensidad. La
concentración de lactato en la célula muscular marcará la fracción que se
convertirá en glucógeno, mientras mayor sea la cantidad más favorecida será la
conversión o resíntesis de glucógeno, a esto se le agrega el bajo pH en la
célula muscular la cual frena la salida del lactato y los diferentes niveles de
intensidad del de ejercicio. En el Ciclo de Cori, el lactato producido del
piruvato, por la reacción de la LDH (lactato deshidrogenasa) es liberado desde
la célula muscular al torrente sanguíneo y transportado al hígado donde es
convertido en glucosa en el interior de hepatocito durante la recuperación; la
glucosa regresa a la sangre para ser usada por el musculo como fuente de
energía y a su vez contribuye en la recuperación del glucógeno hepático, la
cantidad de lactato para esto varía entre el 10 y el 20% del lactato producido
según estudios de (Astrand, 1986 y Krebs, 1963).


3.3. Interrelación de los sistemas energéticos durante el ejercicio.
Los diferentes sistemas energéticos ( anaeróbico aláctico,anaeróbico láctico y aeróbico ) no actúan de forma independiente cuando un individuo esta realizando ejercicio a la máxima intensidad posible. Desde los que realizan actividades cortas, hasta los que realizan actividades de mayor resistencia. Cada uno de los tres sistemas energéticos, están contribuyendo a las necesidades energéticas totales del organismo. Lo que ocurre es que los diferentes tipos de ejercicio y sobre todo con función de la intensidad , predomina un sistema energético sobre los otros.
3.3.1. De acuerdo a la duración del ejercicio
La duración consigue mejorar la capacidad funcional de la población general. La duración de cada periodo puede ir gradualmente aumentando los beneficios se obtienen con secciones acumuladas de 15-20 minutos diarios. Cada sección de ejercicio debe suponer un costo energético de 150 a 400 kcal/ sección. Podemos establecer la duración de de la sección a partir de conocer el kcal gastado de la intensidad establecida en la sección .
3.3.2. De acuerdo a la intensidad del ejercicio
La Intensidad progresiva del esfuerzo desarrollado deberá desarrollarse para alcanzar la duración ideal del test. Los protocolos de valoración aeróbica deberían tener una duración de entre 8 a 12 minutos ya que en un tiempo un poco mas prolongado puede llegar a causar perdida de la concentración del niño.
No hay comentarios:
Publicar un comentario